



美しいパターン形成を、ほ乳類細胞上で再構成する
2019年5月21日
拡散する速度が大きく異なる2つの分子がお互いに反応しながら拡散すると、分子の相互作用によって分子の濃淡の波が発生する。そして、その波によって生き物の形や模様が生み出される。この現象は、1952年にイギリスの数学者であるアラン・チューリングが証明した反応拡散方程式による反応拡散パターンと呼ばれる。生き物の中で自然に発生する一見複雑でとても美しいパターン形成。それを自分の手で培養皿の中に再現することはできるだろうか。
理研BDRの関根亮二研究員(研究当時、再構成生物学研究ユニット、戎家美紀ユニットリーダー)と柴田達夫チームリーダー(フィジカルバイオロジー研究チーム)らは、ほ乳類細胞にNodalとLeftyによる活性化-阻害遺伝子回路を再構成すると反応拡散系によるパターン形成が起こり、それがNodalとLeftyの異なる拡散速度に由来することを明らかにした。本成果は科学誌 Nature Communications に2018年12月21日付で掲載された。
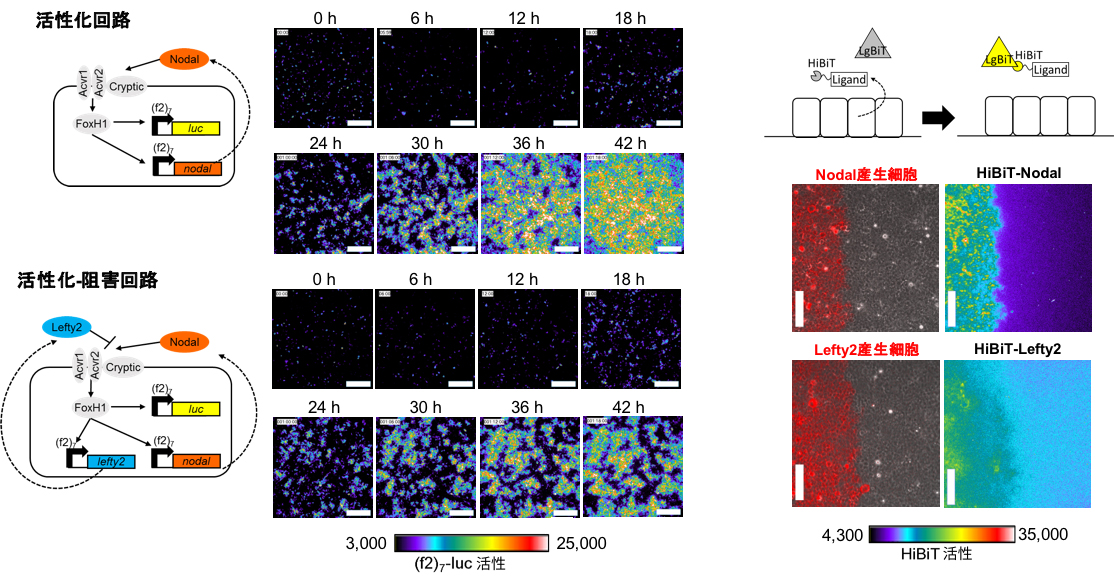
HEK293細胞にNodalによる活性化回路(上)およびNodalとLeftyによる活性化-阻害回路(下)を作成すると、パターン形成が起こった。図に示す分割ルシフェラーゼ(HiBiT)システムで測定すると、細胞外Nodalの拡散範囲は狭く、細胞外Leftyは広い拡散範囲を示した(右)
多細胞生物では、様々な種類の細胞がそれぞれの組織で適切に配置され、その機能を発揮する。この自然に発生するパターン形成を理解するために、細胞上でそのパターンを再構成する研究が続けられている。これまでにいくつかの人工的なパターン形成が報告されているが、まだほ乳類細胞での反応拡散系によるパターン形成の再構成は達成されていなかった。関根らは、ほ乳類の細胞で反応拡散系によるパターン形成を目指した。まず、分泌因子NodalがNodal自身の発現を活性化するような遺伝子回路(活性化回路)をHEK293細胞に作成し、Nodalによるシグナル伝達に応答した遺伝子発現をルシフェラーゼレポーターの発現によってモニターした。この細胞を培養皿に密に播種すると、Nodalレポーター陽性細胞は10%以下でランダムに分布していた。その後、レポーター陽性細胞は徐々に隣接する細胞を活性化していき、42時間後には全ての細胞がレポーター陽性となった。この活性化回路を基盤として、NodalシグナルによってNodalを阻害する分泌因子Leftyも発現するような遺伝子回路(活性化-阻害回路)をHEK293細胞に作成した。この細胞を培養皿に密に播種すると、約18時間後にレポーター陽性細胞の小さなドメインが現れた。その後、ドメインの成長は約30時間後に減速し、約36時間後には明確なレポーター陽性ドメインと陰性ドメインのパターンが形成された。そして、このパターンは約60時間後まで一定に保たれていた。
NodalとLeftyによって、パターンが生じるのはなぜだろうか。ゼブラフィッシュやアフリカツメガエル、マウスの胚においては、NodalとLeftyの拡散速度が異なることが報告されている。今回用いた培養細胞の系でもNodalとLeftyの拡散速度は異なっているかどうかを検討した。細胞外のNodalおよびLeftyを分割ルシフェラーゼで検出するシステムを用いて、それぞれの拡散範囲を測定すると、Nodalはほとんど拡散せず、1つか2つの細胞の範囲に限局していた。一方、Leftyは細胞5〜10個ほどの範囲で遠くなるほどシグナルが減少するような非常に広い分布を示した。NodalにはFinger1ドメイン、Heelドメイン、Finger2ドメイン、Proドメインが存在するが、NodalからFinger1ドメインを取り除くと広く分布するようになった。逆に、このFinger1ドメインをLeftyに移植するとLeftyの分布が狭くなったので、Finger1ドメインが拡散範囲の制御に重要であることがわかった。また、Leftyは細胞の基底側でも頂端側でも同様にぼやけたシグナルとして観察されるのに対し、Nodalは基底側では小さなクラスターを形成しており、Nodalは細胞基底側の培養皿との間の空間に限局して分布している可能性が示された。
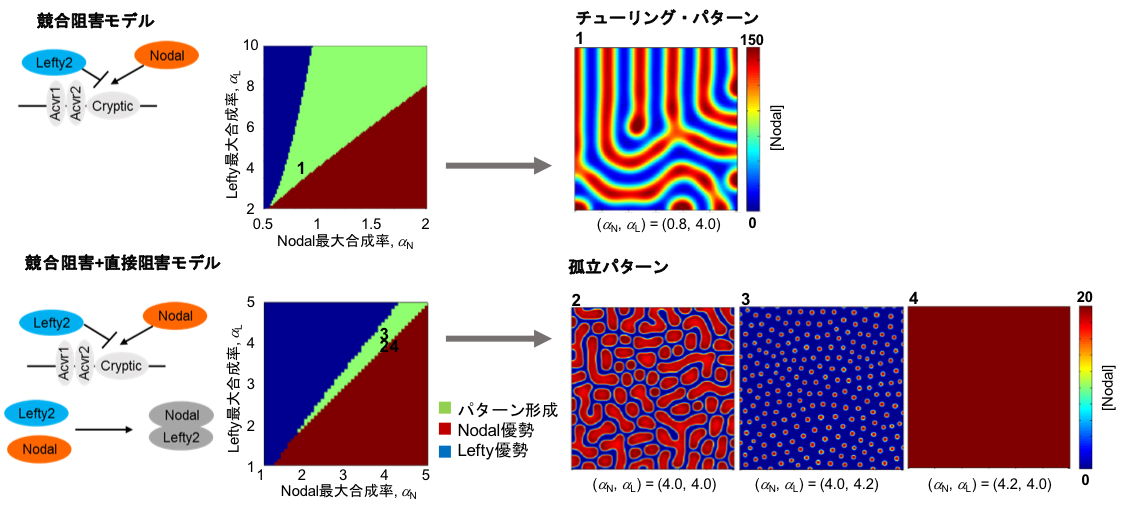
競合阻害モデル(上)と競合阻害+直接阻害モデル(下)から得られるパターン。今回作成した活性化-阻害回路は競合阻害+直接阻害モデルから得られるパターンに近い。
ほ乳類細胞内に作成した活性化-阻害回路をより詳細に理解するために、実際に測定した生化学パラメーターを用いて数理モデルを構築した。LeftyによるNodalの阻害については、受容体への結合を競合的に阻害する、あるいはLeftyがNodalに直接結合して阻害する、という2つのメカニズムの報告がある。そこで「競合阻害モデル」と「競合阻害+直接阻害モデル」の2つのモデルを構築した。競合阻害モデルからは周期的ないわゆる「チューリングパターン」が、競合阻害+直接阻害モデルからはチューリングパターンより規則性の低い「孤立パターン」が得られた。細胞内に作製した活性化-阻害遺伝子回路から生じる実際の細胞のパターンは、細胞播種時のレポーター陽性細胞と陰性細胞の比率を変えると変化したため、初期条件に依存しないチューリングパターンではなく、孤立パターンに近いという可能性が示された。
「ほ乳類の細胞に反応拡散パターンを構築することに世界で初めて成功し、NodalとLeftyはパターン形成の条件を満たすと実証することができました。」と戎家ユニットリーダーは語る。「実際に作ってみると、思ったより全然パターンができなくて驚き苦労するとともに、実際の生き物の美しいパターン形成にはまだ何か秘密があるに違いないと感じています。」
高橋 涼香(BDR・広報グループ)
関連リンク
掲載された論文
