



ヌタウナギとヤツメウナギがHox遺伝子の進化のシナリオを明らかにする
2018年6月18日
動物の体には頭から尾に向かう前後軸があり、それに沿って頭や四肢、尾などが適切な位置に形成される。このような形態形成の位置情報は、Hox遺伝子の発現によって制御されている。Hox遺伝子は哺乳類を含む脊椎動物に限らず、ショウジョウバエなどの節足動物やミミズなどの環形動物を含む、左右相称動物に共通して見出される重要な遺伝子だ。そのため、Hox遺伝子の機能が動物の進化の過程でどのように変遷してきたのかを理解することは、例えばヒトに特徴的な形態がどのように成立したのかを理解することにもつながる。しかし、様々な種を用いたHox遺伝子の解析が進んだ現在でも、その基本的な機能がいつ成立したのかについては、未解明な点が多い。
理研BDRのJuan Pascual-Anaya(フアン パスクアル アナヤ)研究員(形態進化研究チーム、倉谷滋チームリーダー)らは、顎のない脊椎動物である円口類に属するヌタウナギとヤツメウナギのHox遺伝子を解析した。その結果、円口類を含む全ての脊椎動物のHox遺伝子の発現様式は共通しており、現生脊椎動物の共通祖先の段階ですでにHox遺伝子の発現様式が確立していたことを明らかにした。本成果は科学誌Nature Ecology & Evolution に2018年4月2日付で掲載された。
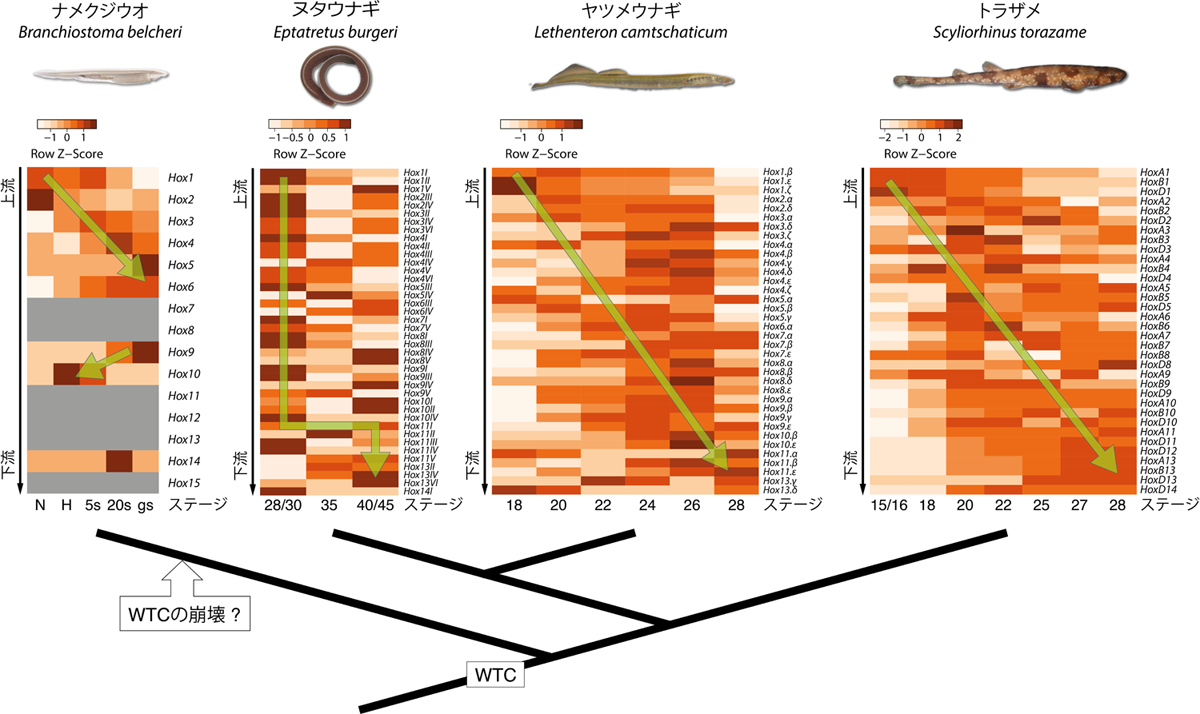
上部はナメクジウオ、ヌタウナギ、ヤツメウナギ、トラザメにおける、Hoxの発生段階ごとの発現強度。縦にHoxが番号順に並べてあり、横軸に発生段階を示す。色が濃い部分でHoxがより強く発現しており、ヌタウナギ、ヤツメウナギ、トラザメの全てで左上から右下に発現が推移していく傾向がわかる。下部は本研究から推定されるHox遺伝子群の進化のシナリオ。共通祖先ではWTC様式のHox遺伝子の発現が成立しており、ナメクジウオでのみその様式が崩壊したのではないかと推定される。
Hox遺伝子群とは、動物の体の前後軸に沿った形態形成に関わるもので、ホメオボックスと呼ばれる共通の配列を持つ13個の遺伝子が、染色体上に一列に並んでクラスターを形成している。このHox遺伝子群の位置と順序には、発生学的に非常に重要な意味がある。まず、Hox遺伝子の染色体上での位置は、遺伝子が発現する体の領域に一致している。例えばHox1は体の前方の領域、Hox6は体の中央部、Hox10は体の尾部というように、前後軸に沿ってそれぞれの領域の形態形成に関与しているのだ。この現象は「空間的共線性」と呼ばれ、これまでに解析されてきたほとんどの動物で見出される。また、Hox遺伝子の発現様式には「時間的共線性」と呼ばれる、遺伝子の位置と発現タイミングの関係性も知られている。これは空間的共線性とは異なり、すべての動物に共通して見られるというものではない。脊椎動物の中で顎のある脊椎動物(顎口類)では、発生が進むに従い、Hox1、次にHox2というように、クラスター内のHox遺伝子が番号順に発現する。それに対し、脊椎動物以外のHox遺伝子はクラスター内でいくつかのグループを作り、そのグループの中でのみ番号順に発現する。前者は「全クラスターの時間的共線性(whole-cluster temporal colinearity:WTC)」、後者は「サブクラスターレベルの時間的共線性(subcluster-level temporal colinearity:STC)」と呼ばれる。では、脊椎動物の共通祖先はWTCとSTC、どちらの様式であったのだろうか。
2つの時間的共線性の成立時期を明らかにするために、Pascual-Anayaらは顎のない脊椎動物である円口類のHox遺伝子について解析を行った。円口類はヤツメウナギとヌタウナギの2系統に分類されるが、近年ヤツメウナギについてはHox遺伝子群の解析が行われており、計43個のHox遺伝子を持つことが報告されていた。そこでまず、Hox遺伝子についての報告が乏しいヌタウナギの解析を行ったところ、ヒト(4クラスター、計39個)とほぼ同数の40個のHox遺伝子が存在することが明らかとなった。また、顎口類のHox遺伝子群がクラスターごとにコンパクトに纏まっているのに対し、ヌタウナギのHox遺伝子は各クラスター内の遺伝子の間が遠く離れていた。これは、ヤツメウナギにも認められる特徴である。次に、ヤツメウナギとヌタウナギのHox遺伝子の発現様式が時間的共線性を示すのかについて、ヤツメウナギとヌタウナギ、顎口類の比較対象としてトラザメの複数の発生段階の胚から得られたHox遺伝子の発現データを解析した。トラザメのHox遺伝子はこれまでの報告どおり明瞭なWTCの発現様式を示した。また、ヤツメウナギもトラザメ同様、明瞭なWTCを示すことがわかった。一方、ヌタウナギではそれほど明瞭な傾向は認められなかったものの、後方のHox遺伝子が発生後期の胚において強く発現している傾向が見られた。以上のことから、円口類を含む全ての脊椎動物はWTCの発現様式による時間的共線性に従ってHox遺伝子を発現することが明らかとなった。つまり、WTCの発現様式は現生脊椎動物の共通祖先の段階ですでに成立していたことになる。
では、WTCの発現様式は脊椎動物より前の、脊索動物の進化の過程で成立したものなのだろうか。この問題を明らかにするために、脊椎動物の最も原始的な祖先に近いと考えられている脊索動物のナメクジウオにおけるHox遺伝子の発現様式について解析を行った。その結果、ナメクジウオではHox1からHox5のみが時間的共線性を示し、WTC様式の時間的共線性は示さないことがわかった。しかし、脊椎動物以外のHox遺伝子が示すSTC様式はHox1からHox3までの最前方のHox遺伝子において認められるものが一般的であり、このナメクジウオで見られる時間的共線性とも異なるものであった。したがって、ナメクジウオで見られるHox1からHox5まで連続する時間的共線性は、WTCが二次的に崩壊した結果ではないかと考えられる。この仮設が正しいとするならば、WTC様式のHox遺伝子の発現様式の起源は、現生の脊索動物の起源まで遡る、より古いものである可能性がある。
「今回、脊椎動物の共通祖先がWTCを示すのみならず、脊索動物の共通祖先までもがWTC様式でHox遺伝子を発現していたらしいことが示唆された。」と倉谷チームリーダーは語る。「今後も、謎の多い生物であるヌタウナギと他の脊椎動物とのゲノム比較が、脊椎動物のゲノム構造の起源と進化についての問題を解く鍵となってゆくだろう。」
高橋 涼香(BDR・広報グループ)
関連リンク
掲載された論文